小鼠iCD1完全诱导培养基 8.29-9.29在丁香通有6折开学促销活动。
链接如下:
http://www.biomart.cn/act/201609_kaixuecuxiao_cellculture_part.htm.htm
iCD1小鼠iPS诱导完全培养基
iCD1完全培养基是一种无血清、无动物源污染、化学成分确定的高效获得小鼠诱导多能干细胞
(iPS细胞)的培养基。该培养基可在无饲养层细胞、不分盘的条件下维持从体细胞向iPS细胞
转化过程中的细胞生长和增殖,同时显著加速iPS细胞诱导进程,极大地提搞iPS细胞的转导
效率。
优点
无血清、无动物源污染、化学成分确定
体系稳定,批次重复性
支持少因子诱导体细胞重编程
极大缩短iPS诱导时间(一周),提高iPS诱导效率(10%)
使用简便,无需添加额外的生长因子或其他添加物
|
产品编号 |
品名 |
规格 |
存储条件 |
|
820210 |
iCD1完全培养基 |
100mL |
-20℃ |
iCD1培养基诱导iPS细胞操作步骤
成纤维细胞导入重编程因子->每天更换iCD1培养基->第7-8天挑取克隆
重编程效果

干细胞特异分子标记物Nanog和内源性Oct4的相对表达量被用于评价细胞整体重编程进程的快慢。
图2a表明,在DeliCell® iCD1培养条件下,Nanog 和内源Oct4的激活明显快于mES和KSR-BN组。
图2b为蛋白水平上验证多能性分子标记物的表达结果图。
结果表明,第8天在原盘出现的重编程克隆其多能性分子标记物Cdh1,SSEA-1和Nanog均有表达。
图2c为将mES和iCD1培养条件下第4天和第8天的培养物提取总蛋白,采用Western-blot检测Nanog
的表达状况,实验结果表明,感染后第8天,在DeliCell® iCD1培养条件下, Nanog表达显著,而
在对照组mES条件下,Nanog则未表达。该结果也进一步证明了iCD1培养条件下重编称过程显著
加快。

图2 从分子水平上证明DeliCell® iCD1培养体系优于其他培养体系。
重编程后期外源因子沉默,是判断完全重编程的一个指标。
图3为采用红色荧光蛋白Ds-Red用于模拟外源转录因子的沉默,将Ds-Red和OKS一起感染细胞,
在DeliCell® iCD1培养条件下,不同时间点,荧光显微镜跟踪观察细胞绿光(指示内源多能性
Oct4的表达)和红光(指示外源因子的表达状况)。
结果显示,随着时间的延长,绿光逐渐增强,红光逐渐减弱,第8天基本不表达红光,
表明重编程后期外源因子的沉默,重编程基本完成。
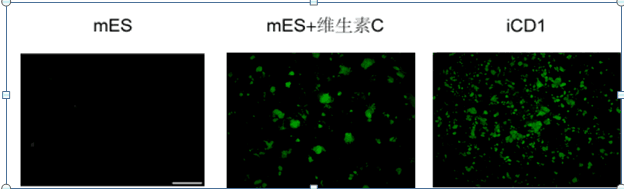
图3 体细胞重编过程示踪

4.2采用iCD1和少因子(OK或OS)诱导小鼠多能干细胞
图4a是在DeliCell® iCD1培养条件下,OK和OS重编程克隆的原始照片和传代照片。
图4b表明OK和OS重编程获得的iPS克隆表达干细胞特异性分子标记。
图4c为Oct4/KLf4和Oct4/Sox2重编程实验结果统计,表中给出了每次实验计算重编程克隆的时间和
统计结果,三次试验结果表明试验具有较好的重复性。图4d为OK和OS克隆注射裸鼠长出的畸胎瘤
组织切边,组织切片中明显的三个胚层的组织分化表明OK和OS克隆具备向三个胚层细胞分化的潜
能。这些数据有力地证明了在iCD1培养条件下,少因子(OK或OS)能够将小鼠成纤维细胞诱导重
编程。

图4 少因子的重编程实验
5.4.3 采用iCD1和单因子Oct4诱导小鼠多能干细胞
MEF细胞用Oct4病毒经过两轮感染后,在iCD1中培养,连续培养30天后,将培养物用胰酶消化
传代种植到饲养层细胞上,用iCD1继续培养,在传代后的第5天左右,发现有绿色重编程克隆出
现,如图5a所示。通过免疫荧光检测发现这些克隆均表达多能性分子标记如Nanog,SSEA-1
和Rex1,结果如图5b所示。
图5c通过实时荧光定量PCR检测多能性marker的表达状况,结果表明这些克隆表达内源Oct4、
Nanog、Rex1、Dppa3、Dnmt3l的水平与标准的胚胎干细胞类似。
图5d是通过实时荧光定量PCR方法分析外源基因的表达状况。OKS感染后4天的Oct4-GFP小鼠
胚胎成纤维细胞作为阳性对照。
结果显示挑取的这些重编程克隆,外源转录因子均已沉默。插入鉴定检测结果表明 (图5e),挑取的
克隆不是因为病毒污染引起。
图5f为Oct4重编程克隆细胞注射囊胚产生的嵌合体小鼠。产生的嵌合体公鼠与白色品系小鼠产下
全黑色小鼠(图5g) ,该结果证明Oct4重编程克隆为生殖系嵌合。
上述结果证明严格证明Oct4重编程细胞与胚胎干细胞类似,从而证明了在iCD1培养条件下,
Oct4可将小鼠胚胎成纤维细胞完全重编程。
1. 培养基相关文献列表:
1. Chen, J. et al. Rational optimization of reprogramming culture conditions for the generation
of induced pluripotent stem cells with ultra-high efficiency and fast kinetics. Cell research 21,
884-894 (2011).
2. Bar-Nur, O. et al. Small molecules facilitate rapid and synchronous iPSC generation.
Nature methods 11, 1170-1176 (2014).
3. Buganim, Y. et al. The developmental potential of iPSCs is greatly influenced by
reprogramming factor selection.Cell stem cell 15, 295-309 (2014).
4. Chen, J. et al. BMPs functionally replace Klf4 and support efficient reprogramming of mouse
fibroblasts byOct4 alone.Cell research 21, 205-212 (2011).
5. Chen, J. et al. Vitamin C modulates TET1 function during somatic cell reprogramming.
Nature genetics 45, 1504-1509 (2013).
6. Papp, B. & Plath, K. Epigenetics of reprogramming to induced pluripotency.
Cell 152, 1324-1343 (2013).
7. Liu, J. et al. The oncogene c-Jun impedes somatic cell reprogramming.
Nature cell biology 17, 856-867 (2015).
8. Chen, J. et al. BMPs functionally replace Klf4 and support efficient reprogramming of
mouse fibroblasts by Oct4 alone.Cell research 21, 205-212 (2011).
9. Wang, Y. et al. Reprogramming of mouse and human somatic cells by high-performance
engineered factors.EMBO reports 12, 373-378 (2011).
10. Ouyang, J. et al. Cyclin-Dependent Kinase-Mediated Sox2 Phosphorylation Enhances the
Ability of Sox2 to Establish the Pluripotent State. The Journal of biological chemistry (2015).
|
|
品牌:Delicell |
|
|
|
||