细胞内遗传物质的稳定受自身与外界多种因素影响,DNA双链断裂(DSB)可以通过细胞呼吸过程中累积的氧化应激自发产生,也可以通过宇宙射线和癌症患者的治疗性辐射外源产生。这些受损DNA打破了细胞稳态,会引起基因突变、染色体畸形,进而导致细胞死亡或恶性转化为肿瘤细胞。衰老、DNA修复、免疫和肿瘤四者是紧密关联的。
维持基因组完整性对于正常的细胞功能至关重要。因此,生物体通过识别DNA损伤位点,激活一系列生化通路,协调DNA复制与转录,使损伤DNA得以修复,维持机体的稳定。DNA损伤修复的研究有助于了解基因突变的机制、衰老和癌变的原因,还可应用于环境致癌因子的检测。表征调控DSB修复的因素和途径对于了解致癌事件期间的诱变过程和开发适当的癌症疗法非常重要。
染色体断裂修复如此重要,那要如何对其不同修复途径进行检测呢?小编带你一起来看!
DSB修复检测系统的原理
DNA损伤的精准修复对于抑制遗传不稳定和肿瘤发生至关重要。
真核生物主要通过两种途径修复:同源重组(HR)和非同源末端连接(NHEJ)。测量途径的典型方法是盒式报告基因检测。
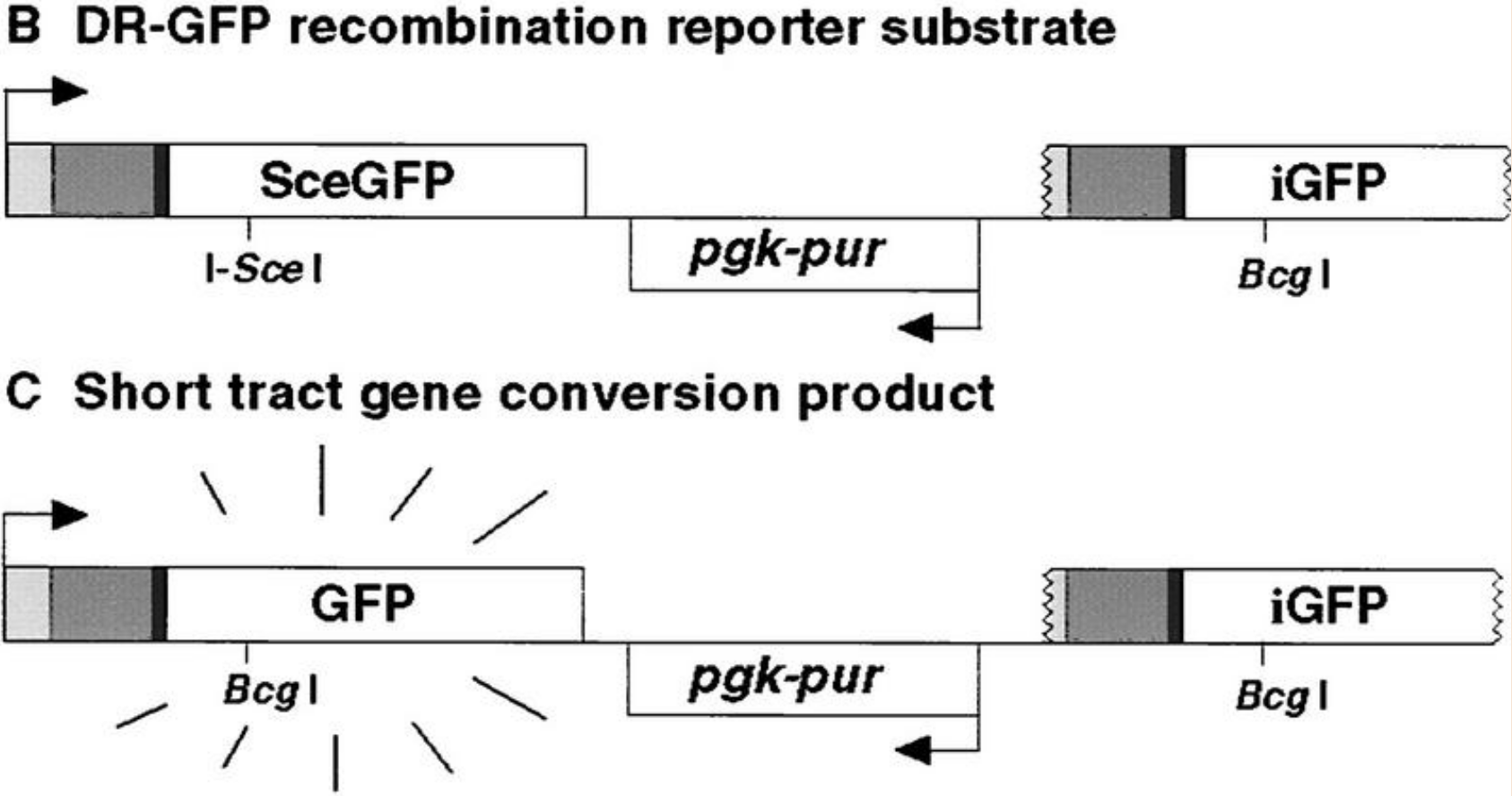
同源重组修复检测系统:
同源重组(HR)检测系统由两个突变的非功能性荧光蛋白SceGFP和iGFP组成,SceGFP和iGFP之间由药物筛选标记嘌呤分离开。
SceGFP作为重组报告基因,序列中插入一个I-Sce I位点使其失活,当加入I-Sce I核酸内切酶时可以引起SceGFP断裂,刺激修复途径激活,以iGFP为供体片段的同源重组修复事件修复断裂的SceGFP进而恢复荧光蛋白的表达。
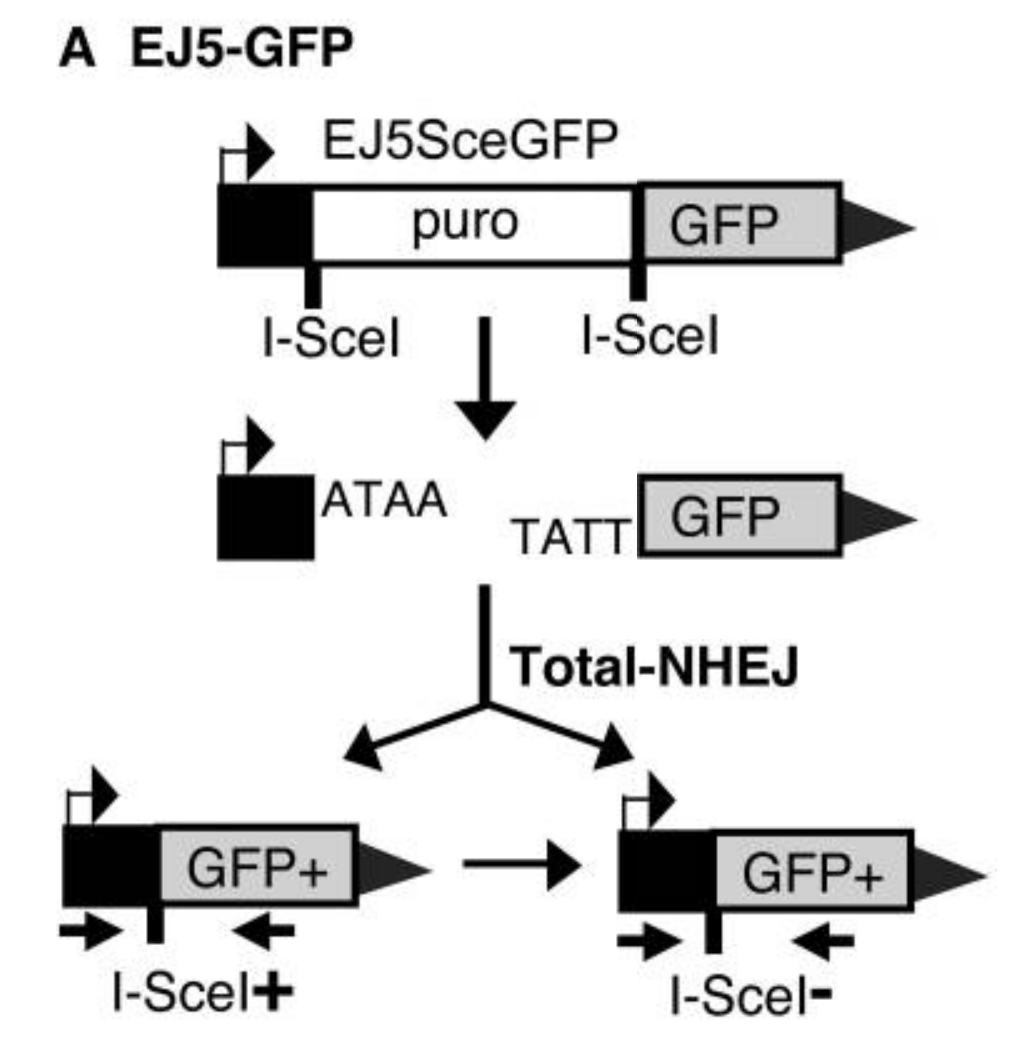
非同源末端连接:
非同源末端连接(NHEJ)检测系统包含一个启动子、嘌呤抗性、GFP;
启动子通过一个puro基因与GFP编码盒分离,puro基因两侧是两个具有相同方向的I-SceI位点,当加入I-Sce I核酸内切酶时,puro基因被切除,NHEJ修复两个I-SceI诱导的DSB,I-SceI识别位点之间的末端连接可恢复GFP+表达。
DSB修复检测系统的应用案例
为研究NORAD如何影响辐射应激期间ESCC细胞的DNA损伤修复反应,在KYSE-150和TE-1细胞中敲低了NORAD。将重组DR-GFP质粒转染到有和没有NORAD敲低的细胞中,以确定NORAD是否调节DNA修复。通过将I-SceI质粒瞬时转染到细胞中来诱导DNA双链断裂。流式细胞术测定显示NORAD敲低导致DSB的同源重组修复减少3-4倍。基于流式细胞术的细胞凋亡测定表明,NORAD敲低提高了照射细胞的凋亡率。证实了NORAD通过调节DNA损伤修复过程来调节ESCC辐射敏感性。

常规实验步骤:
首先,将DNA DSBs报告质粒转染到细胞中,筛选稳定表达DR-GFP的细胞,并瞬时转染另一个表达I-Sce I核酸内切酶的质粒。
ISceI转染2-4天后,使用流式细胞术评估GFP阳性细胞的比例,并计算修复效率。
吉凯基因可以提供用于染色体断裂修复检测系统的质粒:
HR同源重组修复的pDRGFP 质粒;
NHEJ非同源末端修复的EJ5GFP质粒(EJ5-GFP检测多类NHEJ事件,因此可以被认为是总NHEJ的测定);
I-Sce I核酸内切酶的质粒。
如果您有其他检测系统需求,也欢迎咨询。